Os tecidos fundamentais são aqueles originados do meristema fundamental e desempenham diversas funções no corpo da planta.
Parênquima:
O parênquima é o tecido mais abundante no corpo da planta e pode ser encontrado em todos os órgãos vegetais adultos. É constituído por células vivas, fisiologicamente ativas e originadas do tecido fundamental. Caracteristicamente possui espaços intercelulares (meatos), cujo tamanho varia conforme a função do tecido.
As células parenquimáticas possuem forma, tamanho e conteúdo variados. São geralmente grandes, poliédricas, com parede celular primária delgada (fina), vacúolo grande e conteúdo conforme a função na planta. No citoplasma podem ocorrer leucoplastos, cromoplastos ou cloroplastos e no vacúolo é comum a ocorrência de inclusões sólidas (cristais) e antocianina. A parede celular pode em alguns casos se lignificar.
De acordo com a função ou o conteúdo o parênquima pode se diferenciar em:
· Parênquima de preenchimento: encontrado no córtex de caules, raízes, na medula dos caules, pericarpos de frutos entre outros. As células são isodiamétricas com espaços intercelulares pequenos.
Parênquima medular em escapo
floral de Hemerocalis sp.
· Parênquima clorofiliano (clorênquima): ocorre em órgãos que realizam fotossíntese, em especial a folha. É rico em cloroplastos e conforme a forma, a disposição das células e o volume dos espaços intercelulares pode ser denominado de parênquima clorofiliano propriamente dito, o parênquima paliçádico (células em formato de palitos) e o parênquima lacunoso; os dois últimos são encontrados principalmente nas folhas.
Corte transversal da lâmina foliar
Corte transversal da lâmina foliar mostrando parênquima clorofiliano.
· Parênquima de reserva: encontrado em órgãos de reserva, as células acumulam amido, lipídios e proteínas.
· Parênquima aequífero: ocorre em plantas suculentas (cactáceas, euforbiáceas, crassuláceas) sendo especializado no armazenamento de água. Suas células são relativamente grandes, com enorme vacúolo e fina camada de citoplasma.
· Parênquima aerífero (aerênquima): comum em plantas aquáticas caracteriza-se pela presença de espaços intercelulares volumosos (lacuna) e interconectados dando origem a uma fase gasosa contínua dentro da planta. O aerênquima é um tecido relativamente forte, apesar de ser bastante leve, e a forma de suas células varia bastante, desde isodiamétricas até braciformes.
Corte da folha submesa de Cabomba caroliniana (Eudicotiledônea) mostrando o aerênquima.
Bona et al (2004)
Bona et al (2004)
As células do parênquima ocorrem também associadas aos tecidos condutores, recebendo o nome de parênquima do floema (associado ao floema), ou lenhoso (associado ao xilema). Em muitas partes da planta ocorrem células parenquimáticas especializadas no transporte rápido e a curta distância de materiais- as células de transferência-, as quais apresentam parede labiríntica com inúmeras protusões voltadas para a face interna e acompanhadas de plasmalema, o que implica em significativo aumento em área facilitando o transporte.
As células do parênquima retém, mesmo quando maduras, a capacidade de se dividir, retornando a atividade meristemática e desempenhando papel importante no processo de regeneração e cicatrização.
Colênquima:
É um tecido formado de células vivas, relativamente alongadas, de paredes primárias celulósicas, espessadas, relacionado com a sustentação de regiões jovens, em crescimento, no corpo do vegetal. O tecido origina-se, principalmente, do meristema fundamental. É um tecido semelhante ao parênquima, ambos apresentam protoplasto vivo, podem possuir cloroplastos e até mesmo reassumir características meristemáticas, voltando a apresentar divisões celulares. A diferença entre estes dois tecidos está, principalmente, no fato do colênquima, geralmente, apresentar suas células mais alongadas e com paredes mais espessas que as células do parênquima.
Característica e função:
A estrutura da parede celular é a principal característica do colênquima. As paredes são primárias, celulósicas com pontoações primordiais e, geralmente, o espessamento das paredes é irregular. Essas paredes apresentam grandes quantidades de substâncias pécticas, que são altamente hidrofílicas. Assim, as paredes das células do colênquima retêm uma grande quantidade de água (60% do seu peso), o que as torna extremamente plásticas capazes de acompanhar o crescimento das células.
O colênquima é um tecido adaptado para a sustentação de regiões jovens do vegetal em intenso crescimento. O espessamento das paredes de suas células inicia-se bem cedo, antes mesmo da célula completar o seu crescimento, mas a plasticidade destas paredes possibilita o crescimento do órgão, até que este atinja a sua maturidade.
O grau de espessamento da parede parece estar relacionado com as necessidades do vegetal. Por exemplo, em plantas sujeita à ação de ventos fortes, o espessamento das paredes do colênquima inicia-se precocemente e é maior, do que aquele observado em plantas que crescem sob condições mais amenas. O colênquima maduro é um tecido forte e flexível, formado por células alongadas reunidas em feixes. Por outro lado, nas regiões periféricas de órgãos jovens que são fotossintetizantes o colênquima pode ser clorofilado. Como essas regiões jovens são tenras e, portanto, mais facilmente atacadas pela herbivoria, a cicatrização e regeneração celular pode ser conseguida, devido à capacidade do colênquima de reassumir a atividade meristemática, voltando a apresentar divisões celulares, promovendo assim a cicatrização das regiões lesadas.
Classificação do colênquima:
De acordo com a distribuição do espessamento nas paredes celulares, observado em secções transversais do tecido, podemos reconhecer quatro tipos de colênquima:
Quando as paredes são mais espessas nos pontos de encontro entre três ou mais células, como por exemplo, no pecíolo de Begonia (begônia), caule de Ficus (figueira), de Coleus e de Curcubita (aboboreira).
Colênquima angular; 1. depósito interno à parede celular; 2. Lumen celular.
Colênquima lamelar:
As células mostram um maior espessamento nas paredes tangenciais interna e externa, como o visto no caule jovem de Sambucus (sabugueiro).
Colênquima lacunar:
Quando o tecido apresenta espaços intercelulares e os espessamentos de parede primária ocorrem nas paredes celulares que limitam estes espaços.
Colênquima anelar ou anular:
Quando as paredes celulares apresentam um espessamento mais uniforme, ficando o lume celular circular em secção transversal.
Relação forma-função:
O colênquima, devido a plasticidade e capacidade de alongar-se, adapta-se à sustentação das folhas e caules em crescimento. O grau de espessamento parece estar relacionado com as necessidades da planta pois em espécies sujeitas a ação de agentes externos, como por exemplo, ventos fortes, o espessamento das paredes celulares inicia-se precocemente e torna-se mais acentuado do que o espessamento de espécies sob condições amenas.
Por outro lado, as regiões jovens, geralmente são fotossintetizantes, necessitando, portanto, de tecidos que permitam a entrada da luz, como e o caso do colênquima. Ainda estas regiões, geralmente são tenras e, portanto mais facilmente atacadas por herbívoros, além de microorganismos, havendo necessidade de cicatrização e regeneração celular; estes fenômenos são conseguidos devido a capacidade do colênquima de reassumir a atividade meristemática. Em regiões mais velhas da planta, o colênquima pode se transformar em esclerênquima, pela deposição de paredes secundárias lignificadas.

Colênquima próximo à epiderme da nervura principal (100x)
http://www.labot.uff.br/laminario/Araceae-Dieffenbachia/
Colênquima angular próximo à epiderme da nervura foliar (aumento 100x).
http://www.labot.uff.br/laminario/Apocynaceae-Nerium-oleander/
Esclerênquima:
É caracterizado pela elasticidade das suas paredes que podem ser deformadas por tensão e pressão reassumindo forma e tamanho original quando as forças desaparecem.
É tecido de sustentação de órgão que maduros onde o crescimento longitudinal já cessou.
Vivas ou mortas na maturidade com parede secundária espessa e lignificada conferindo rigidez e pouca permeabilidade.
O esclerênquima é um tecido de sustentação, com parede secundária espessa e uniforme, possuindo cerca de 35% de lignina, o que lhe fornece um revestimento estável, evitando ataques químicos, físicos ou biológicos. É encontrado em vários órgãos e pode formar faixas ou calotas ao redor dos tecidos vasculares, fornecendo proteção e sustentação. Há basicamente dois tipos celulares no esclerênquima: as fibras e as esclereides.
Fibras:
As fibras são células longas e largas, com paredes secundárias espessas e lignificadas, suas extremidades são afiladas, lume reduzido (55 cm Rami, 6 cm Cânhamo) e possuem a função de sustentar partes do vegetal que não se alongam mais.
Podem estar associadas ao xilema (xilemáticas) ou não (extraxilemáticas) situadas no córtex ou no mesofilo.
Quando associadas ao floema podem acumular grãos de amido.
Aspecto geral das fibras (www.uoguelp.ca/botany/courses
Já as esclereides são células isoladas ou em grupos esparsos, distribuídas por todo o sistema fundamental da planta. Possuem paredes secundárias espessas, muito lignificadas, com numerosas pontuações simples.






Tipos de esclereides:
Braquiesclereídes ou células pétreas: são isodiamétricas, ocorrendo, por exemplo, na polpa de Pyrus (pêra) e no marmelo, onde aparecem em grupos entre as células parenquimáticas.
Braquiesclereíde da polpa de Pyrus sp. (www.cas.muohio.edu/ ~meicenrd/ANATOMY )
Macroesclereídes: são células alongadas ou colunares distribuídas em paliçadas e podem ser encontradas no envoltório externo (testa) das sementes das leguminosas, por exemplo, em Pisum (ervilha) e Phaseolus (feijão).
Macroesclereíde do pecíolo de Camellia sp. Foto de Castro, N. M.
Osteoesclereídes: esclereídes alongadas, com as extremidades alargadas, lembrando a forma de um osso, como as esclereídes observadas sob a epiderme (tegmen) da semente das leguminosas (células em ampulheta).
Macroesclereíde do pecíolo de Camellia sp. Foto de Castro, N. M.
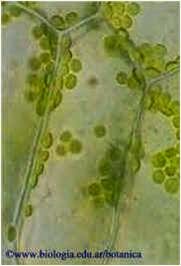
Astroesclereídes: com a forma de uma estrela, com as ramificações partindo de um ponto mais ou menos central. São comumente encontradas nas folhas de Nymphaea sp (lírio d'água).
Astroesclereídes no aerênquima do caule de Numphoides sp. (www.biologia.edu.ar/)
Tricoesclereídes: esclereídes alongadas, semelhante à tricomas, ramificados ou não, como vistas nas folhas de Olea deliciosa (oliveira) e nas folhas de Musa sp (bananeira).
Tricoesclereídes da folha de Musa. Foto de Curtis, Lersten e Nowak.
Referências:
APPEZZATO B. G. & GUERREIRO-CARMELLO S. M. Anatomia Vegetal. Viçosa: Imprensa Universitária UFV, 2003.
CUTTER, E. Anatomia vegetal parte 1 – Células e Tecidos. São Paulo: Roca, 1986. 304p.
ESAU, K. Anatomia das plantas com sementes. São Paulo. Edgard Blücher/USP,1974. 293 p.
BONA, C; BOEGER M. R. & SANTOS, G. O. Guia Ilustrado de Anatomia Vegetal. Ribeirão Preto: Holos Editora, 2004.





















.png)




















